


























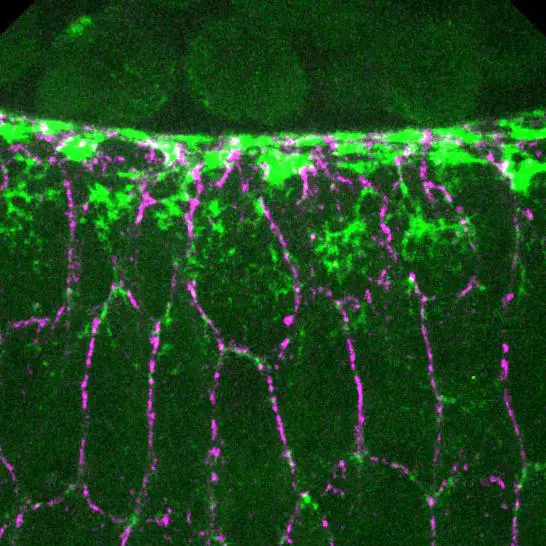
Zooming in on a wave of tissue invagination
Understanding how adhesion and contractility drive a wave of tissue invagination from a subcellular perspective.
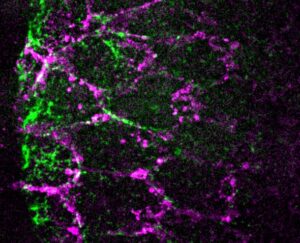
Sanjay Karki from Thomas Lecuit’s team reports that serotonin signaling regulates actomyosin contractility during embryonic axis morphogenesis in insect and bird.
IBDM Marseille inspires young minds: engaging primary school children on childhood cancer (“Contre le cancer, j’apporte ma pierre”) and interacting with high school students through immersive experiences (DECLICS).

Internal Seminar by Swedha Sidharthan
Join us on 08/06/2023 at 12:30 in Amphi 12 for an exciting talk by Swedha Sidharthan from our Team!
Several awards for IBDM members !
Congrats to Aziz Moqrich, Sophie Chauvet and Stefan Harmansa.

Formation of polarized contractile interfaces by self-organized Toll-8/Cirl GPCR asymmetry
Jules Lavalou and Qiyan Mao from Thomas Lecuit’s team report that Toll-8 controls myosin-II planar polarity in Drosophila embryos and wing discs via a physical interaction with the GPCR Cirl/latrophilin.
Embryo development like a stadium wave
In a recent study appeared on the international journal Nature, Thomas Lecuit and his colleagues at the Institut de Biologie du Développement de Marseille describe how tissue shape changes are self-organized.

